Глюконеогенез и его обходные пути
Эта тема входит в мой курс по биохимии
Глюконеогенез — это синтез глюкозы из других соединений. Давайте сначала поговорим о том, почему наш организм синтезирует глюкозу. Ведь это делается не в одну химическую реакцию: для этого существует целый метаболический путь — серия реакций. Так что это запарное дело.
Организм получает много глюкозы в ходе пищеварения. Большая часть полисахаридов и дисахаридов включает в себя глюкозу. Какие-то только из неё и состоят. После переваривания глюкоза попадает в кровь, а оттуда в клетки. Клетки используют глюкозу по своему усмотрению. Например, в мышечной ткани и печени она запасается в виде гликогена. Между приёмами пищи эти запасы разрушаются. Мышцы используют гликоген для того, чтобы обеспечить энергией себя. Но вот печень выбрасывает глюкозу в кровь, где она используется другими клетками. Нервные клетки и эритроциты не могут получать энергию из других источников: жиров и белков. С нервными клетками есть оговорки, но о них в другой раз. Главное понять, что глюкоза — это лучшее топливо для нервной ткани и эритроцитов.
Получается, что мозг и эритроциты целиком зависят от запасов гликогена в печени. Но этот запас не бесконечен! Его хватает, дай Бог, на сутки. Это только первая проблема. Есть ещё и другая: гликоген синтезируется после приёма пищи.
А теперь представьте, что мы не покушали денёк. Или решили пробежать марафон. А вдруг эти две ситуации совпали? Тогда наш мозг должен отключиться. Но этого не происходит. Нам тяжело, но мы продолжаем функционировать. Всё из-за того, что глюкоза синтезируется в ходе глюконеогенеза. Потом она попадает в кровь, а оттуда в ткани и органы. Большая часть глюкозы отправится в нашу святая святых — мозг.
Глюконеогенез работает при голодании и длительных физических нагрузках. Его основная задача — обеспечить мозг и эритроциты глюкозой.
Надеюсь, что вы убедились в важности глюконеогенеза. Да? Тогда можно говорить о нём подробнее.
Глюконеогенез
Я боюсь запутать тебя, если скажу, что глюконеогенез — это обратный гликолизу процесс. Но этого хватит для того, чтобы понять его суть. В ходе разбора мы поймём, что это не так (или не совсем так). При гликолизе мы разрушали глюкозу до пирувата или лактата. В зависимости от типа гликолиза: аэробного или анаэробного. Почему бы не повернуть гликолиз обратно? Так из двух молекул пирувата можно будет получить глюкозу.
Клетка примерно так и делает, но с одним большим НО. Гликолиз — это необратимый процесс. Если мы посмотрим на него, то увидим такое…

Первая, третья и десятая реакция не могут идти в обратную сторону. Поэтому весь гликолиз — необратимый процесс. Но почему эти реакции необратимы?
В ходе первой синтезируется глюкозо-6-фосфат, а в ходе третьей — фруктозо-1,6-бисфосфат. У этих реакций есть кое-что общее. В них гидролизуется макроэргическая связь ( отмечена «~») в молекуле АТФ. Что приводит к высвобождению большого количества энергии — эти реакции экзотермические. Небольшая часть энергии тратится на реакцию фосфорилирования — при этом нет образования новой макроэргической связи. Большая часть рассеивается в виде тепла. Если мы захотим повернуть реакцию в другую сторону, то нам нужно затратить такое же количество энергии. В клеточных условиях такое сделать невозможно.
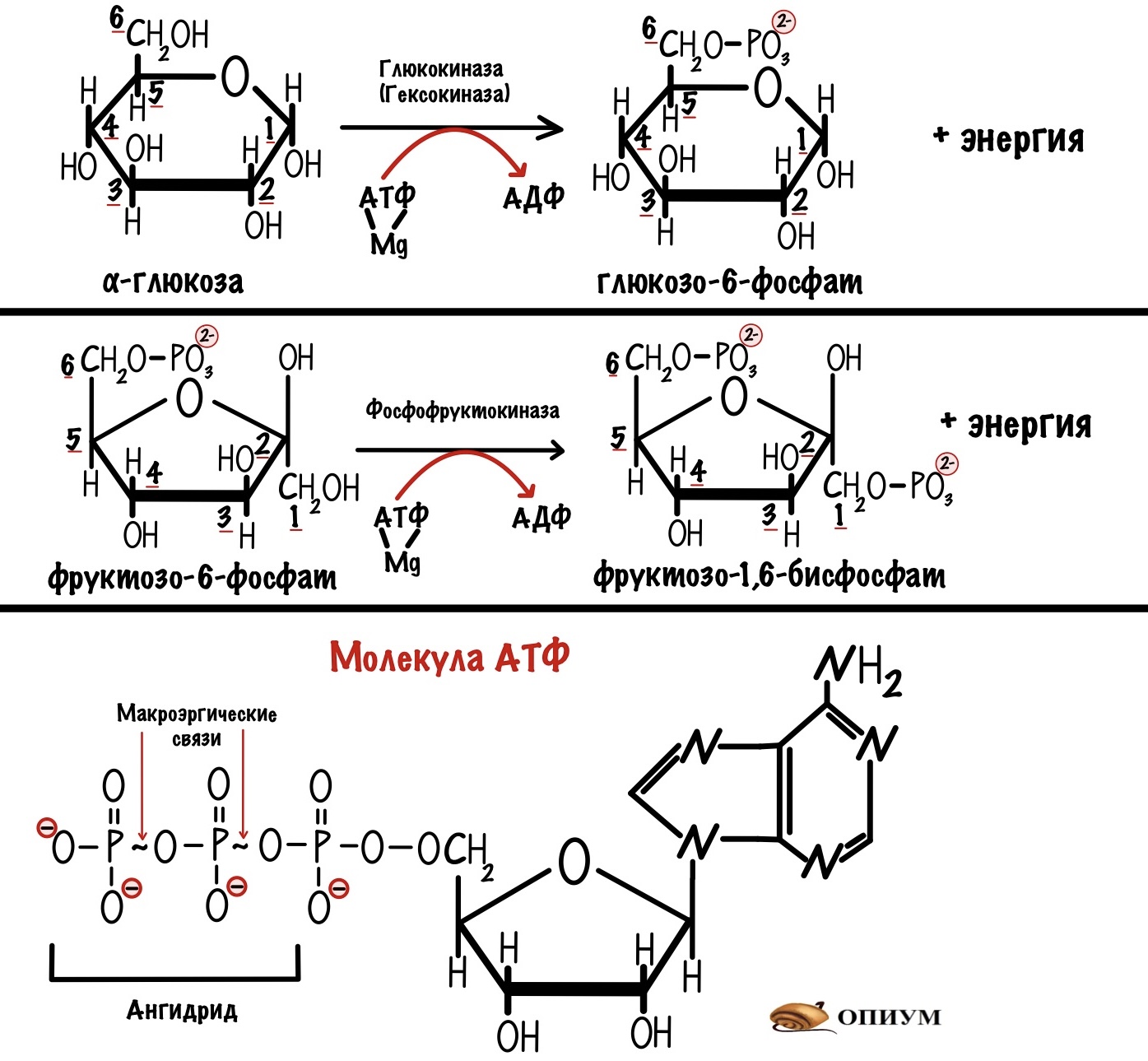
В десятой реакции из фосфоенолпирувата образуется пируват и молекула АТФ. Это реакция субстратного фосфорилирования. С двух сторон в этом уравнении есть макроэргические связи: слева в молекуле фосфоенолпирувата, а справа в молекуле АТФ. Кажется, что она должна быть обратимой. Но нет! От фосфоенолпирувата отщепляется фосфорная группа — образуется енольная форма пирувата. Енольная форма нестабильна, поэтому она быстро переходит в кетонную форму. Кетонная форма более стабильна, поэтому при клеточных условиях мы не можем повести реакцию в другую сторону. Так что эта реакция тоже необратима.

Выходит, что глюконеогенез не может быть обратным гликолизом. Это просто невозможно. В клеточных условиях нам не повернуть эти три реакции в другую сторону. Но, как говорится, умный в гору не пойдёт, умный гору обойдёт.
Клетке нужно обойти только три этих необратимых реакции. Ведь остальные семь — обратимые. Клетка воспользуется ими для синтеза глюкозы. Поэтому суть глюконеогенеза — обойти три необратимые реакции гликолиза. Клетка делает это через создание обходных путей. Но на это способны не все клетки, а только клетки определённых органов. Глюконеогенез идёт в печени, корковом веществе почек и слизистой тонкого кишечника. Только в клетках этих органов есть ферменты, которые проводят реакции по обходным путям. Наиболее активно глюконеогенез протекает в печени.
Суть глюконеогенеза — обойти три необратимые реакции гликолиза.
Глюконеогенез идёт в печени, корковом веществе почек и слизистой тонкого кишечника.
Всего обходных пути три, как и необратимых реакций гликолиза. Давайте посмотрим на гликолиз и глюконеогенез в сравнении:
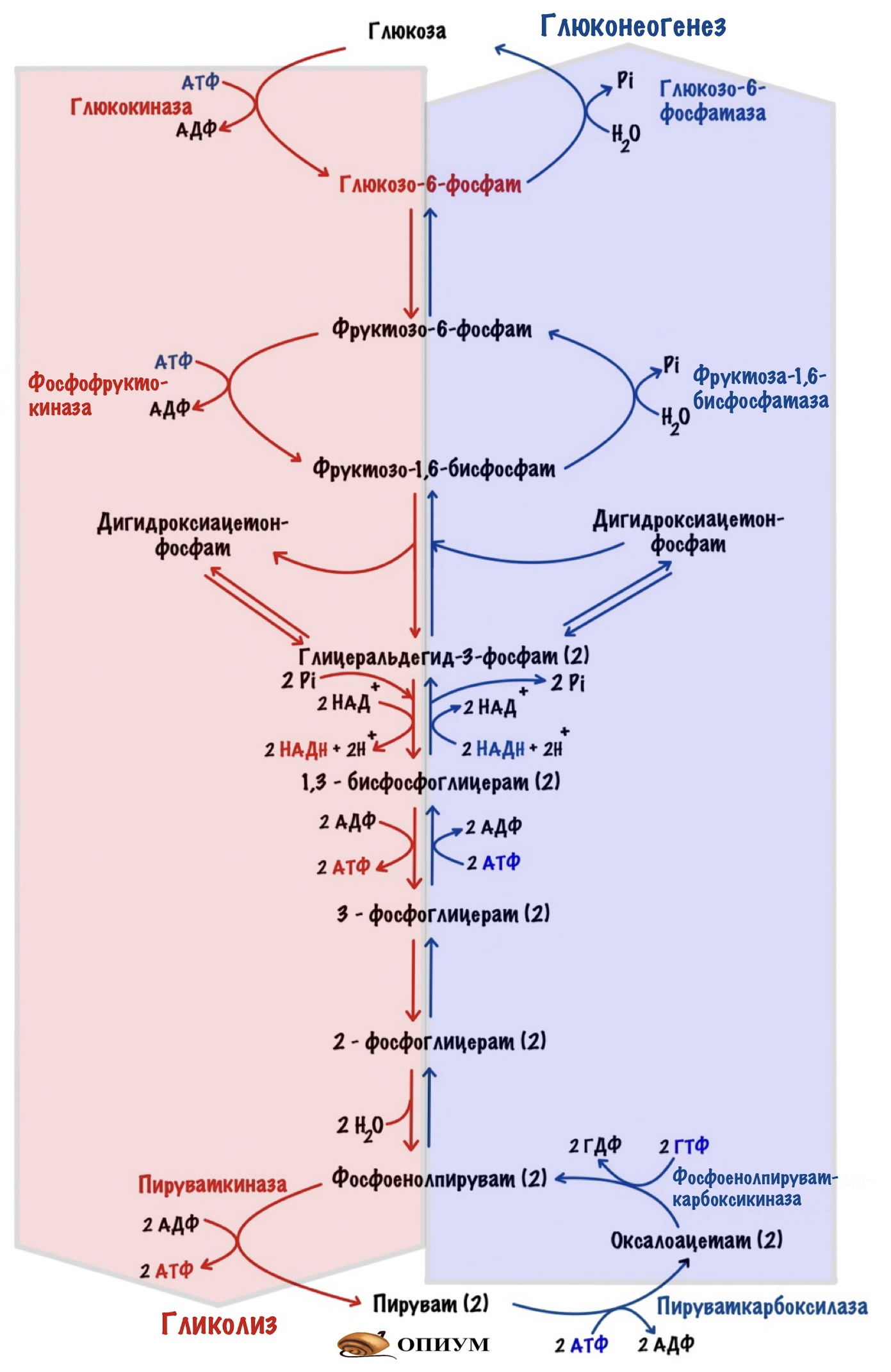
Мы начнём разбирать глюконеогенез с пирувата. Он будет нашим первым субстратом. Чуть ниже посмотрим на другие субстраты глюконеогенеза.
Обходной путь 1. Образование фосфоенолпирувата
Пируват, который образовался в ходе гликолиза был транспортирован в митохондрию. Нам повезло, потому что первый фермент обходного пути тоже находится в матриксе митохондрии. Этот фермент — митохондриальная пируваткарбокилаза.
Пируваткарбоксилаза присоединяет к пирувату CO2 — образуется оксалоацетат.
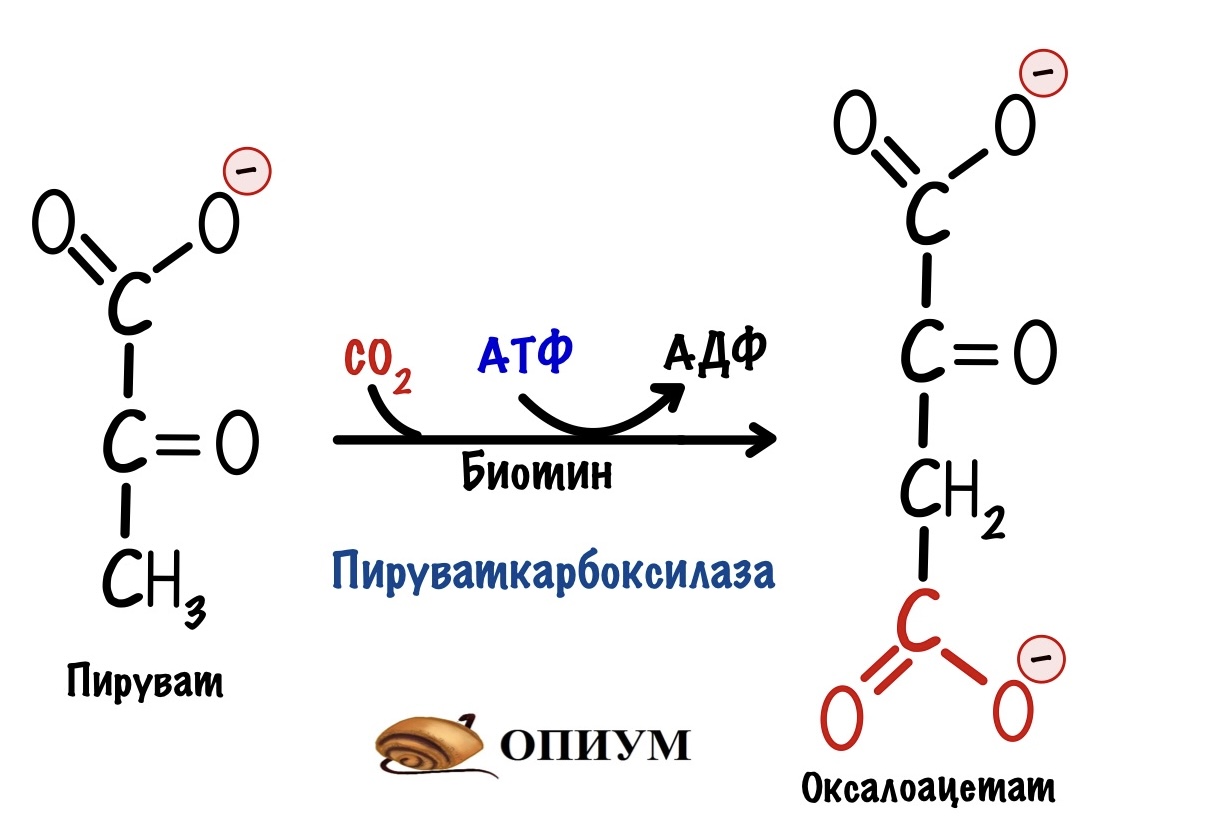
Для этой реакции нужны молекула АТФ и кофермент — биотин. АТФ тратится на активацию бикарбонатного иона, так как у нас в организме большая часть CO2 растворена. Из бикарбонатного иона образуется CO2, который присоединяется к биотину. Биотин переносит CO2 на молекулу пирувата, клетка получает оксалоацетат.
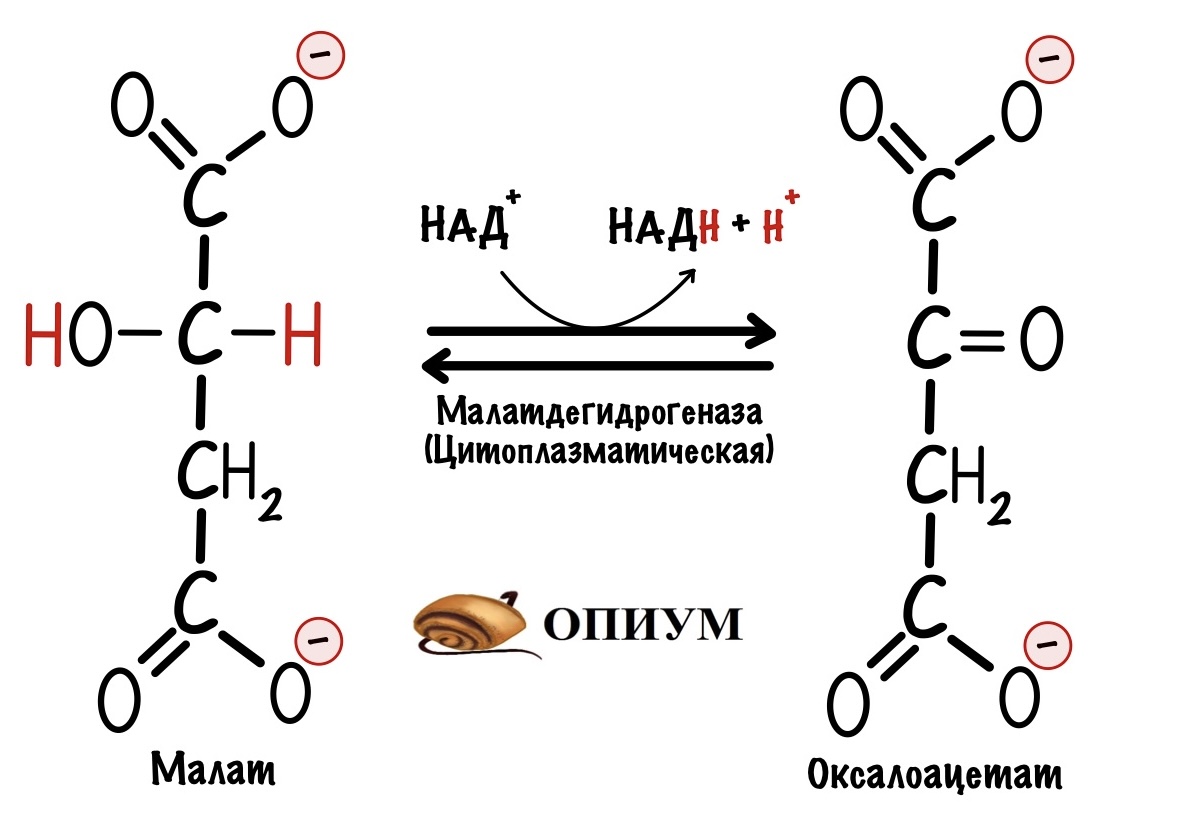
Дальше нужно перенести оксалоацетат в цитоплазму клетки — там находятся семь ферментов, которые катализируют обратимые реакции гликолиза. Для оксалоацетата во внутренней мембране митохондрий нет переносчика. Но есть для малата. Поэтому сначала оксалоацетат превращается в малат. Катализатор — митохондриальная малатдегидрогеназа. Она присоединяет два протона и два электрона к молекуле оксалоацетата. Кофермент реакции — НАДH. Так образуется малат.
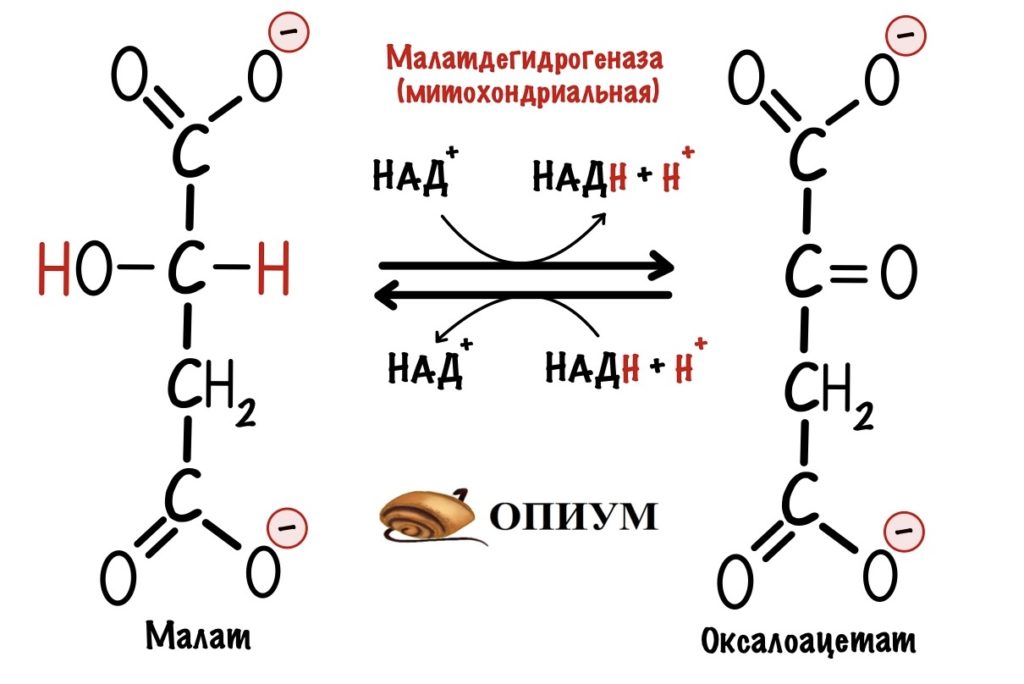
Митохондриальная малатдегидрогеназа обычно катализирует обратную реакцию — превращение малата в оксалоацетат. Так было в малат-аспартатном челноке, но почему здесь всё по-другому? Дело в том, что в митохондрии находится избыток НАДH, а не НАД+. Поэтому реакция идёт в другую сторону. Избыток НАДH обеспечивает окисление жирных кислот — запускается при голодании и/или физической нагрузке.
Далее малат транспортируется через внутреннюю мембрану с помощью белка-переносчика. В наружной мембране есть поры, поэтому дальше он влетает в цитоплазму.

В цитоплазме клетки малат превращается в оксалоацетат с помощью цитоплазматической малатдегидрогеназы. При этом образуется оксалоацетат и НАДH. Эта реакция полезна, так как в шестой реакции для образования глицеральдегид-3-фосфата нужен НАДH. Так митохондрия передаёт избыток протонов и электронов на нужды цитоплазмы — всё это через образование малата.
Теперь нужно превратить оксалоацетат в фосфоенолпируват. Для этого нужен второй фермент этого обходного пути — фосфоенолпируваткарбоксикиназа. Он получил название по обратной реакции. Кроме фермента, нужен источник энергии и фосфатной группы — здесь используется ГТФ. Образуется фосфоенолпируват, ГДФ и CO2. Так клетка обходит десятую реакцию гликолиза.

Пока что глюконеогенез дорогой процесс. Мы затратили молекулу АТФ и ГТФ на образование фосфоенолпирувата. А эта молекула содержит только три атома углерода. Для образования глюкозы нужно образовать два фосфоенолпирувата. То есть общие затраты энергии — 2 молекулы АТФ и 2 ГТФ.
Затраты энергии в первом обходном пути: 2 АТФ и 2 ГТФ.
Дальше идут обратимые реакции до образования фруктозо-1,6-бисфосфата (третья реакция гликолиза). В ходе седьмой реакции мы затратим ещё две молекулы АТФ на образование двух 1,3-бисфосфоглицератов.
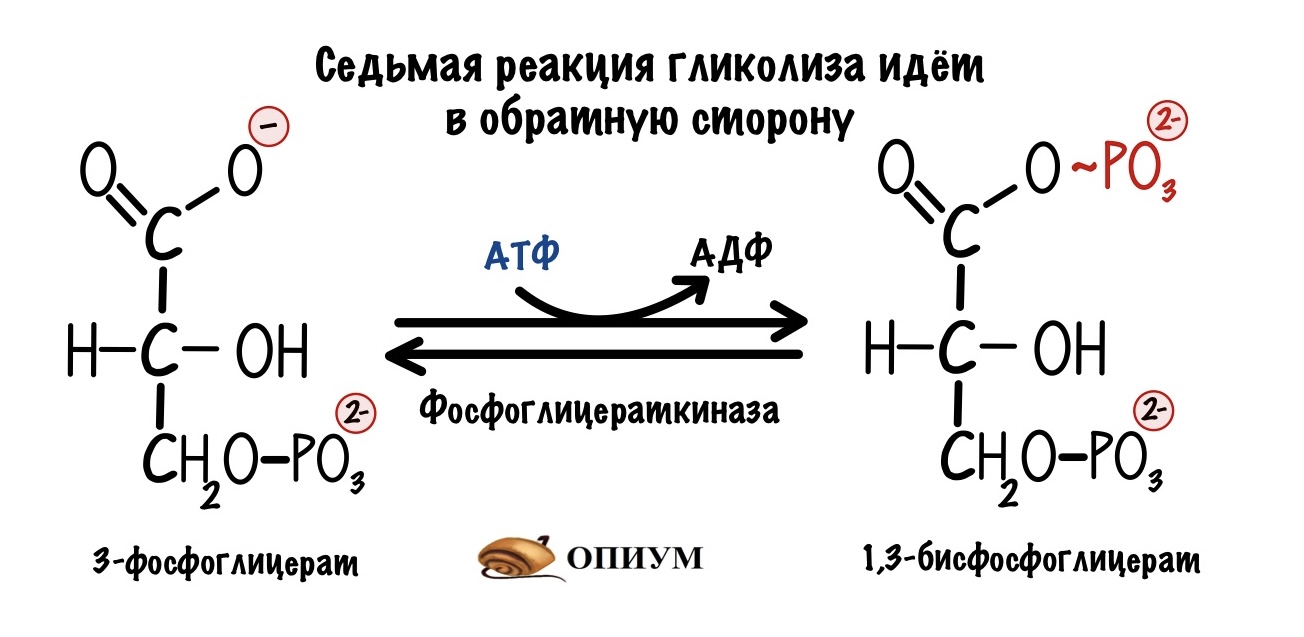
Общие затраты энергии — 4 молекулы АТФ и 2 молекулы ГТФ. Дальше клетка не будет тратиться на синтез глюкозы, так что это общие затраты на весь глюконеогенез.
На синтез одной молекулы глюкозы нужно: 4 АТФ, 2 ГТФ и 2 пирувата.
Дороговато
Хоть этот процесс и дорогой, но других вариантов у организма нет. Нельзя же бросить мозг и эритроциты — без них тело долго не проживёт.
Обходной путь 2. Образование фруктозо-6-фосфата
Здесь всё проще. В этот обходной путь вступает фруктозо-1,6-бисфосфат. Фермент фруктозо-1,6-бисфосфатаза гидролизует одну эфирную связь в этой молекуле. Образуется фруктозо-6-фосфат и неорганический фосфат.
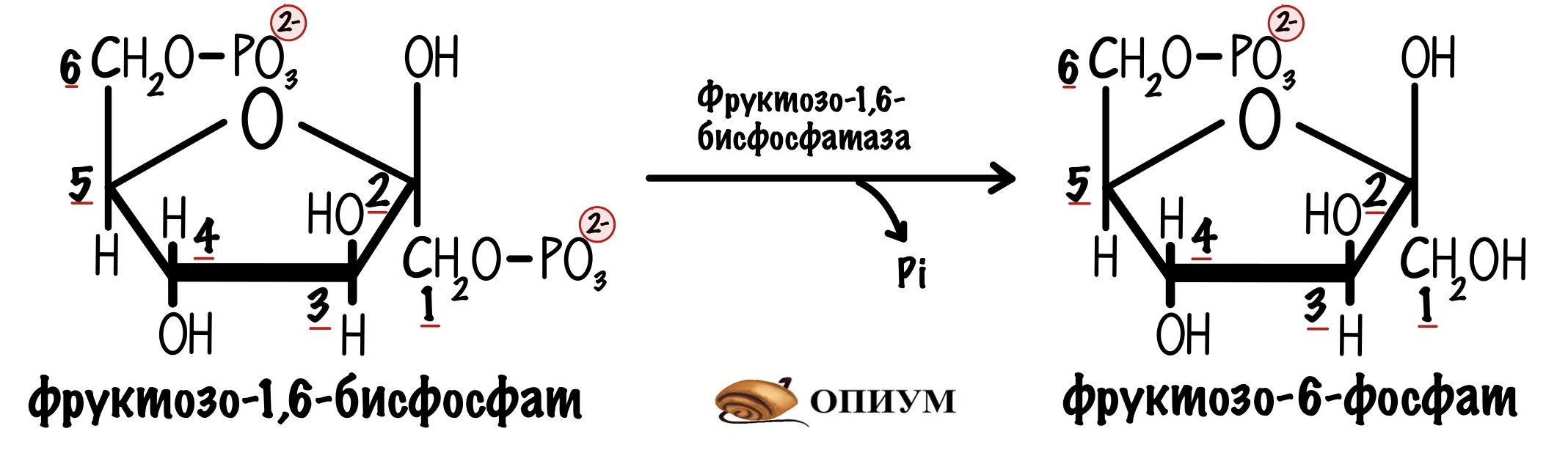
Фруктозо-6-фосфат вступает во вторую реакцию гликолиза — образование глюкозо-6-фосфата. Эта реакция обратима, поэтому не интересует нас.
Обходной путь 3. Образование глюкозы
От глюкозо-6-фосфата отщепляется фосфатная группа. Катализирует реакцию глюкозо-6-фосфатаза.
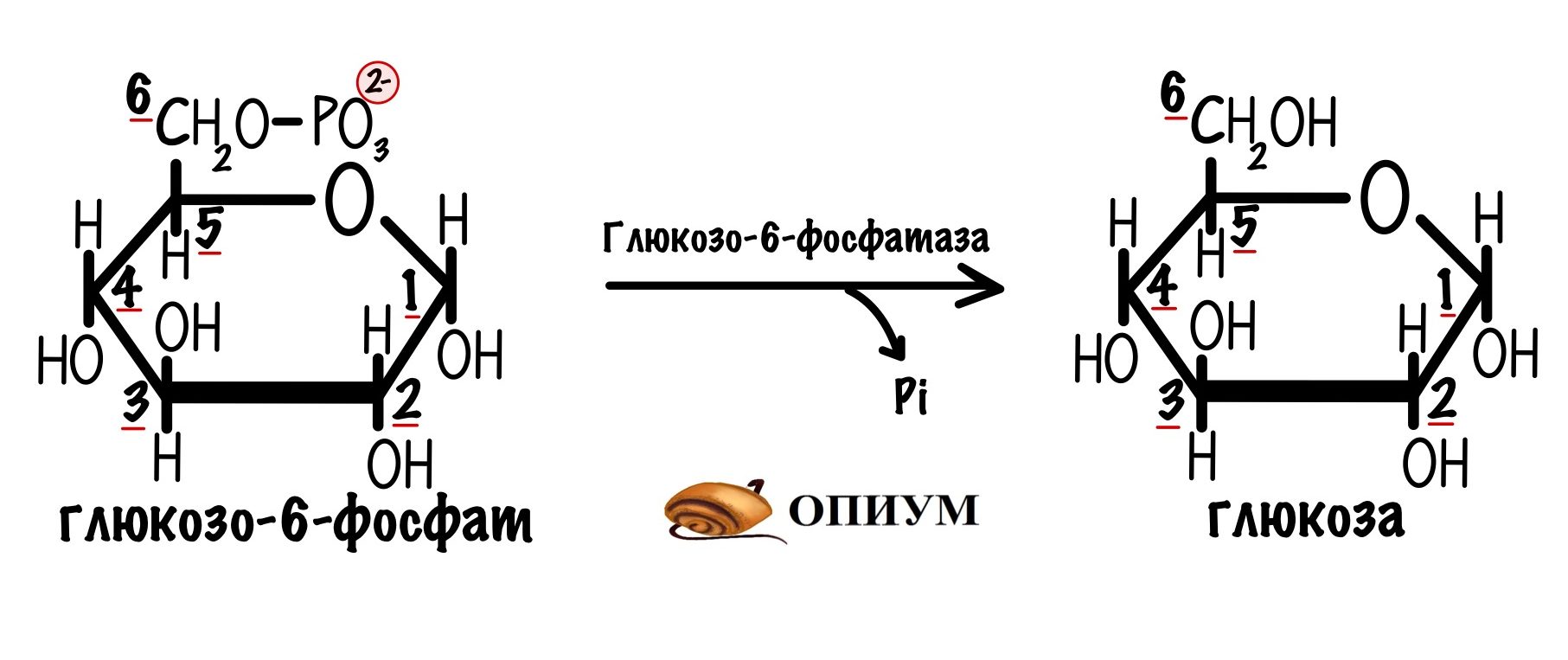
Но этот фермент не может находиться в цитоплазме. Почему? Тогда клетка не смогла бы выполнять свои функции. Представьте, что глюкоза попадает в печень. Она сразу же фосфорилируется с помощью глюкокиназы. Так клетка добивается двух вещей:
- Увеличивает реакционную способность глюкозы;
- Не даёт глюкозе выбраться из клетки. Потому что для глюкозо-6-фосфата нет белков переносчиков в цитоплазме. А вот для обычной глюкозы есть — глюкозный транспортёр 2 типа (ГЛЮТ-2).
Если мы засунем в эту систему фермент глюкозо-6-фосфатазу, то она будет постоянно превращать глюкозо-6-фосфат в глюкозу. А та будет вылетать из клетки. Получается, что вся система работает в холостую. Поэтому глюкозо-6-фосфатаза не работает в цитоплазме, а находится в эндоплазматическом ретикулуме (ЭПР). Он отделён от цитоплазмы биологической мембраной.
В мембране ЭПР есть белки переносчики для глюкозо-6-фосфата. Он переносится в ЭПР, где дефосфорилируется глюкозо-6-фосфатазой. Образуется глюкоза и неорганический фосфат. Они переносятся в цитоплазму, опять же с помощью своих белков переносчиков. А дальше глюкоза попадает в кровь через глюкозный транспортёр второго типа.
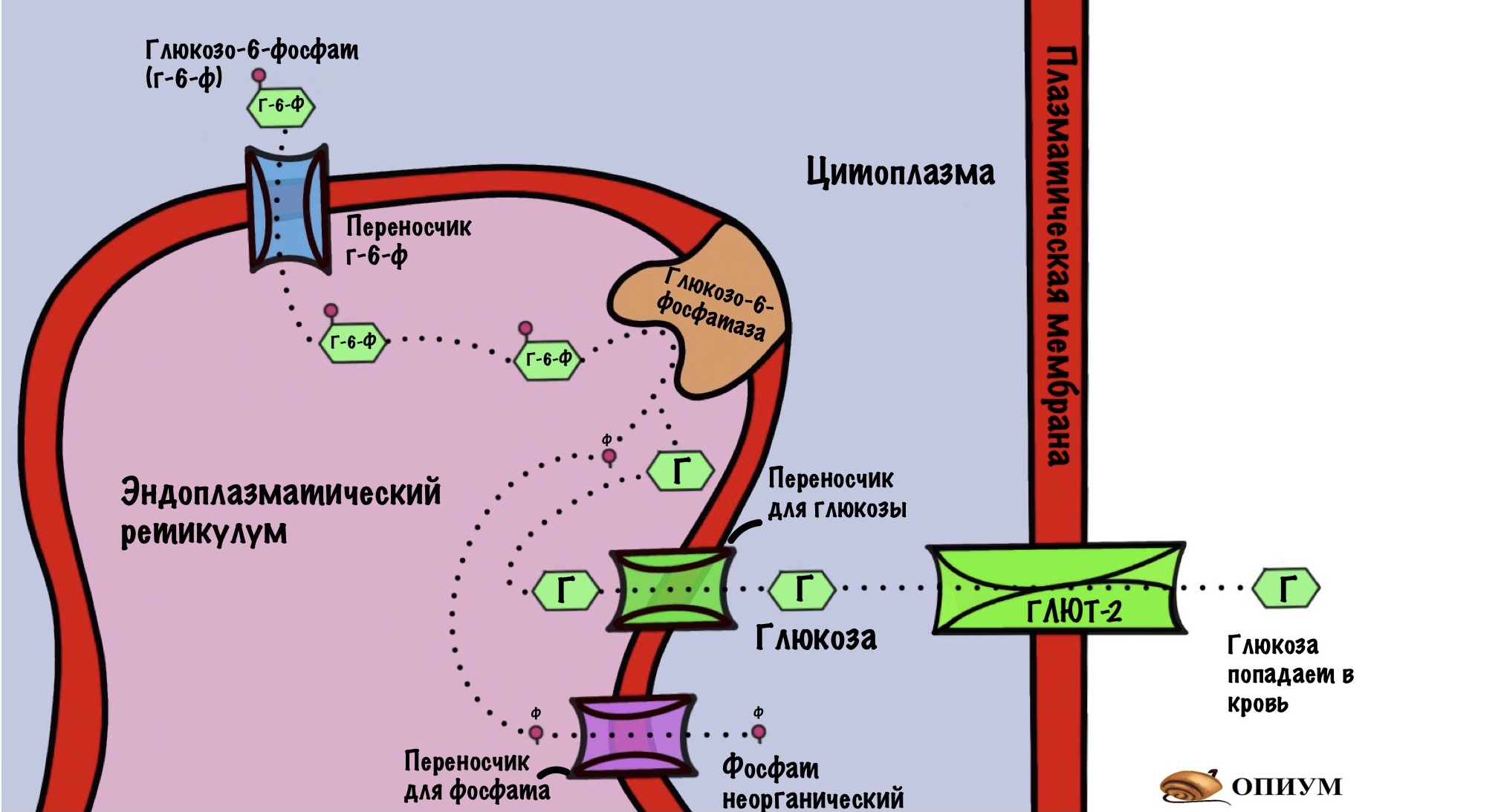
Вот и весь глюконеогенез. Держите общую картинку.
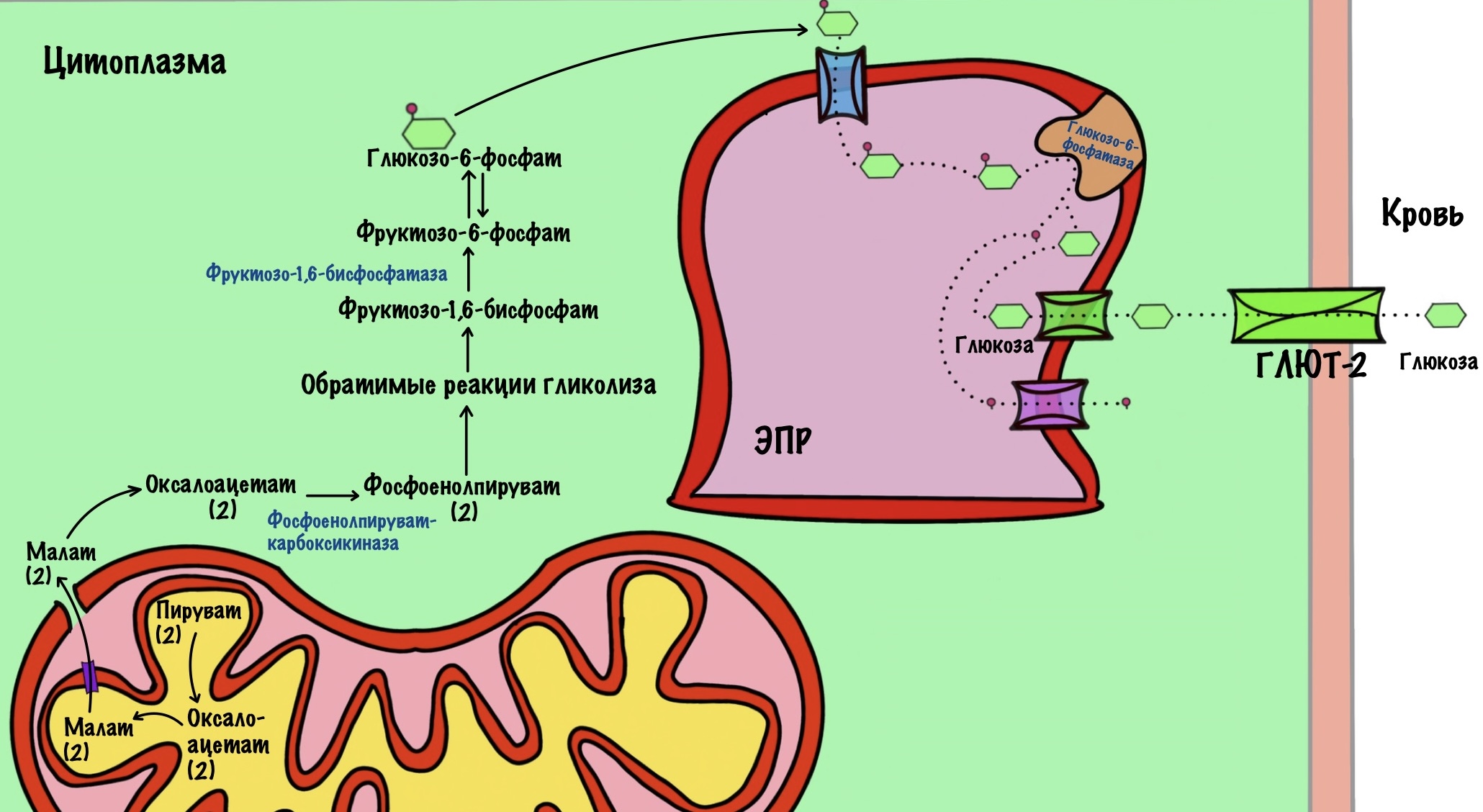
Субстраты глюконеогенеза
Мы разобрали всё на примере пирувата. Но ведь и другие молекулы могут вступать в глюконеогенез, если превратить их в пируват или оксалоацетат. Так лактат образуется из пирувата в ходе анаэробного гликолиза, такая реакция идёт в мышцах и эритроцитах:
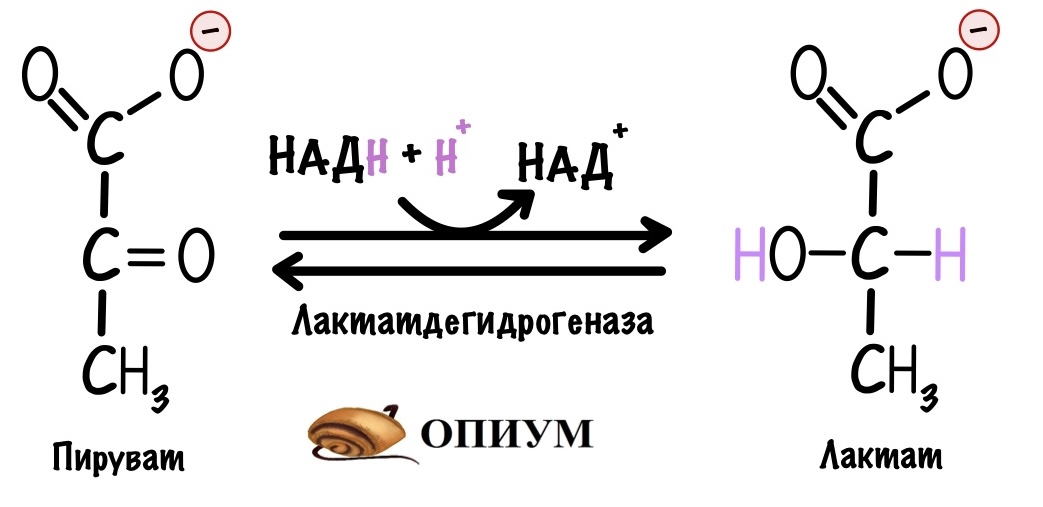
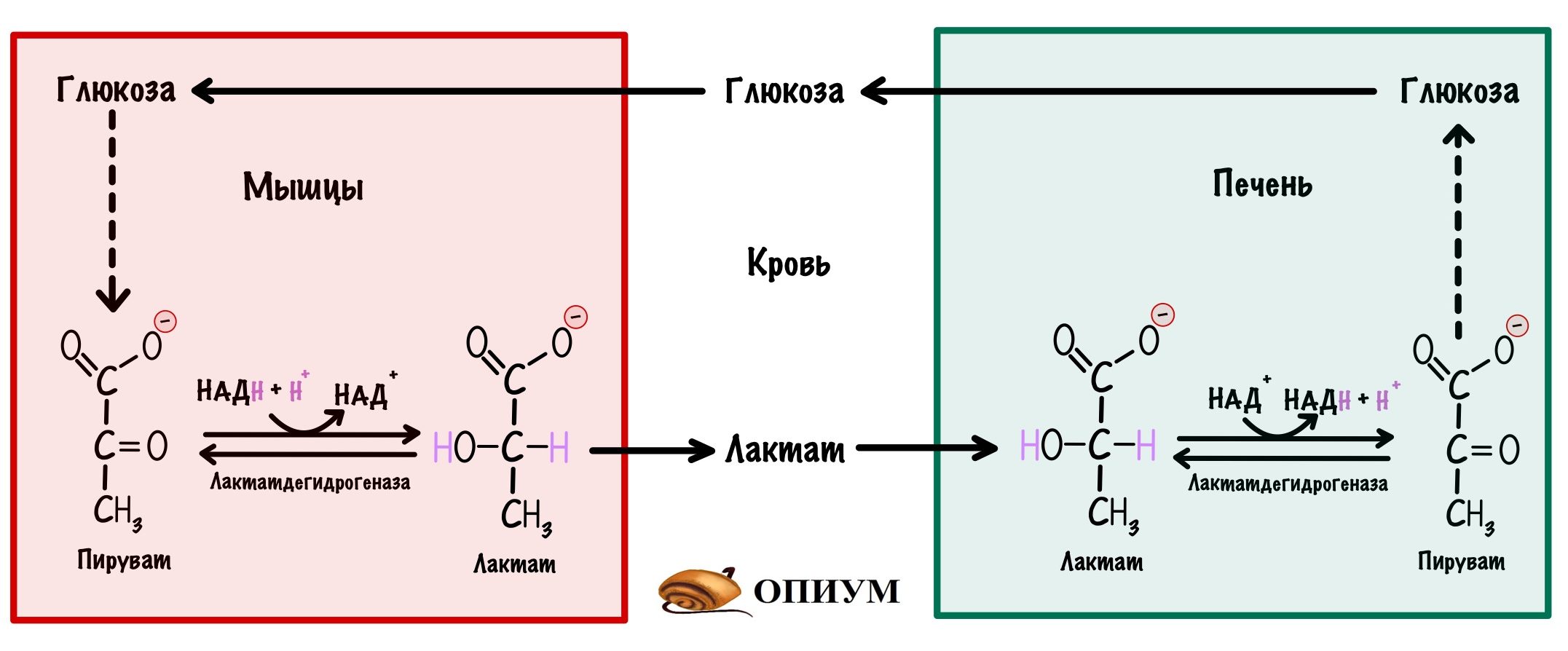
Мышцам и эритроцитам нечего делать с лактатом, поэтому они отправляют его в кровь. Там лактат циркулирует, пока не попадёт в печень или почки. Здесь он может превратиться в пируват, который вступит в глюконеогенез. Катализирует реакцию лактатдегидрогеназа, но реакция идёт в другую сторону. Всё это из-за того, что в печени больше НАД+, чем НАДH.
Так из лактата образуется глюкоза, которая снова может попасть в мышцы. У циркуляции лактата между мышцами и печенью есть название — цикл Кори.
Оксалоацетат образуется в ходе цикла трикарбоновых кислот (ЦТК), поэтому все эти кислоты можно назвать субстратами для глюконеогенеза.
Ещё в глюконеогенез вступают аминокислоты — все, кроме лизина и лейцина. Самый простой пример — аланин. Из него с помощью реакции трансаминирования можно получить пируват.
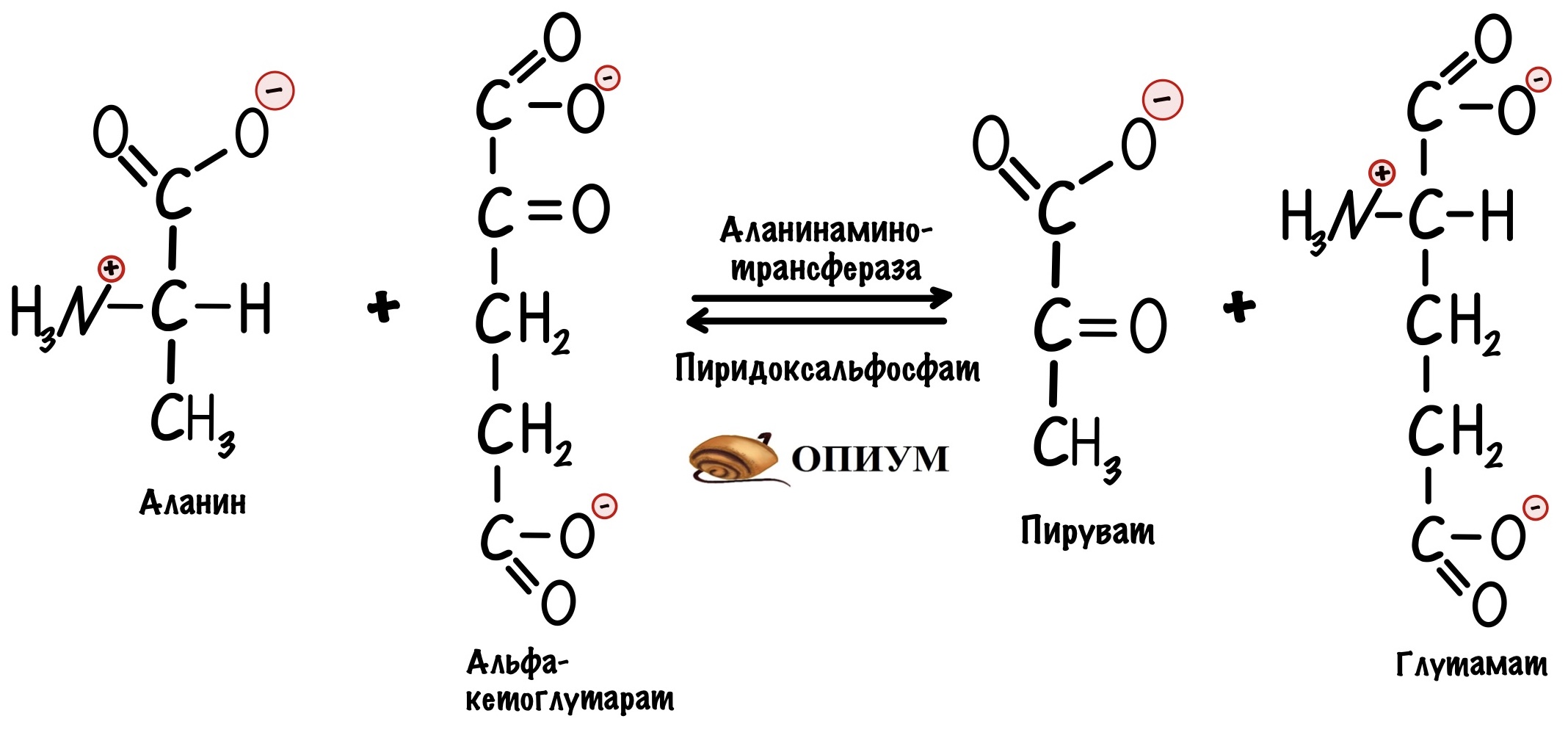
А вот синтез оксалоацетата из аспартата:
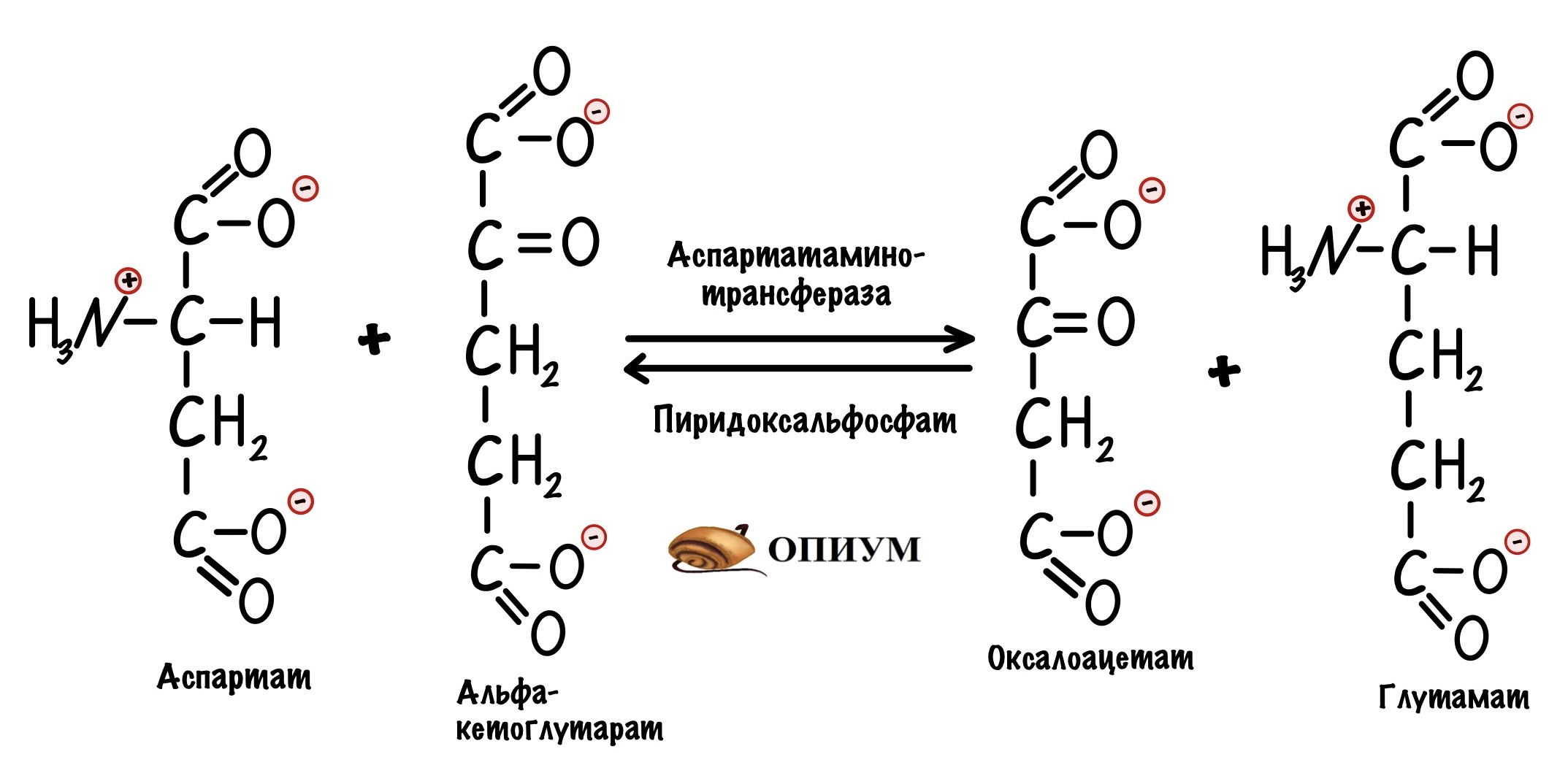
С другими аминокислотами реакции сложнее, но не будем закапываться в этом.
Жирные кислоты не вступают в глюконеогенез, но вот глицерин, который образуется в ходе гидролиза жиров, может вступать. Правда немного по другому механизму. Глицерин фосфорилируется с помощью глицеролкиназы. Образуется глицерол-3-фосфат.
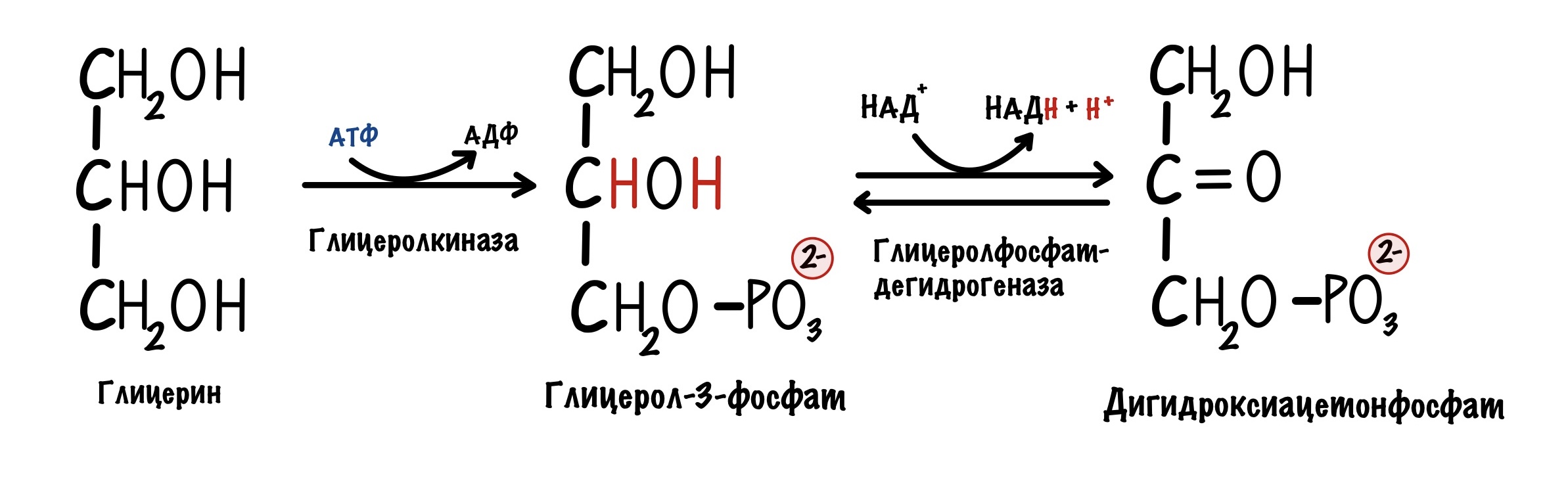
Далее от глицерол-3-фосфата отщепляется два атома водорода у второго углерода. Катализирует реакцию глицеролфосфатдегидрогеназа. Образуется наш старый знакомый — дигидроксиацетонфосфат. Продукт четвёртой реакции гликолиза. Дигидроксиацетонфосфат будет участвовать в образовании фруктозо-1,6-бисфосфата.
Таким путём глюконеогенез избавляет организм от избытка глицерина, который образуется при распаде жиров.
Субстраты глюконеогенеза: пируват, лактат, глицерин, аминокислоты и кислоты из ЦТК.
Регуляцию глюконеогенеза и гликолиза можно почитать здесь.
